
Polarization Sensitivity: A Strong and Weak Sense
by Meghan Rouillard
February 2011
This article appears in the February 4, 2011 issue of Executive Intelligence Review and is reprinted with permission.
Research Report
- The Extended Sensorium: Overview
- Synesthesia: Beyond the Five Senses—Oyang Teng
- Helen Keller: Mind over Instrumentation—Meghan Rouillard
- Following the Beat of a Different Drummer—Peter Martinson
- Polarization Sensitivity: a Strong and Weak Sense—Meghan Rouillard
- What is Circularly Polarized Light?—Jason Ross
- Insects and Infrared—Oyang Teng
- Magnetoreception—Benjamin Deniston
- Unheard Melodies: Electric and Magnetic Senses in Humans—Sky Shields
- The Sounds of a Cosmic Chorus—Aaron Halevy
Bows and Bees
Our eyes are able to distinguish polarized from non-polarized light[1] only very faintly without the aid of other visual devices. When visible, this appears as a small yellow and blue bowtie image in the center of the visual field, called Haidinger’s brush—try staring at the white screen of your laptop, while tilting your head slowly to the side, to see it. Otherwise, our eyes require polarized filters to distinguish it. That is not to say that we don’t see polarized light without them; we just typically don’t see it as something that stands out against light that is not polarized. We will return to the human biological polarization sense later, but for now, let us compare the first known cases of human and animal navigation using polarized light.
Many years ago, it is thought that the Vikings used a crystalline “sunstone” to determine the location of the Sun on very cloudy days, for navigational purposes. Crystals are known to polarize light, and to produced polarized light of different colors. Surely this could have been used to infer the position of a light source, but it is thought that certain kinds of crystals, such as quartz, tourmaline, or cordeirite, which could have served the purpose of a sunstone, could also have indicated the angle of incoming sunlight through changing color and brightness, to perhaps indicate the position of the Sun, even indirectly, through polarized sunlight patterns in the sky.[2]
Animals have been found to operate based on a similar principle, though, of course, they do not use crystals as supplemental instruments. The capability to distinguish polarized light by some birds, insects,[3] and a few sea creatures, is more developed than our own. Early in this study, the polarization sense was surmised to be used by bees, which can additionally sense the Earth’s magnetic field, and are known to dance based on gravitational cues, and the position of the Sun.[4] The Sun sense and the polarization sense were found to be closely related. The bees’ dance, based on knowing the location of the Sun, is used to give directions to other bees, to indicate where a distant food source may lie, and they have been found to use this dance when a food source is 100 or more meters away from the hive. In this dance, the Sun’s position is the key reference point. This dance is called the “waggle waggle dance” (by humans, of course).
 Courtesy of J. Tautz and M. Kleinhenz, Beegroup Wurzburg |
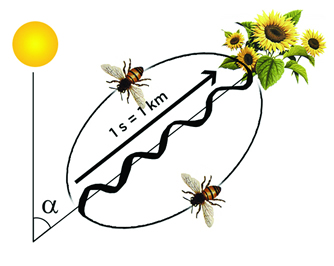 LPAC/Chris Jadatz |
| The honeybees’ waggle waggle dance. In the diagram, s indicates the time during which the waggle part of the dance takes place, and the distance to the food source. The angle alpha, an angle on the honeycombs between the vertical direction and the waggle part of the dance, indicates the angle between the Sun’s position and the direction of flight to be taken. Another variation of the dance occurs when it is done on a horizontal plane, but the orientation towards the Sun is still necessary. | |
These dances were studied by an Austrian ethologist (student of animal behavior), Karl von Frisch. He says his main discoveries were made in 1944, but were not accepted until decades later. He noticed that when he prevented the bees from seeing the Sun’s light, or when they were exposed to diffuse light, their dance became disoriented, but when exposed to even only a very small portion of the blue sky, they would resume the dance as though the Sun were in view. This led him to assume that the bees were responding to the polarization of light from the Sun.[5] Here we quote von Frisch’s account of his discovery in his 1973 Nobel Lecture:[6]
There can be no doubt that the Sun’s position is decisive for the direction of their dancing.... But there was one big puzzle. To prevent excessive heating during most of the experiments, a protective roof was installed over the observation hive. The dancers were unable to see the Sun. Nevertheless their dance was usually correct. Orientation by heat rays, by penetrating radiation, as well as other explanations that seemed possible had to be discarded—until I noticed that a view of the blue sky is the same as a view of the Sun. When clouds passed over the section of the sky visible to the bees, disoriented dances immediately resulted. Therefore they must have been able to read the Sun’s position from the blue sky. The direction of vibration of polarized blue light[7] differs in relation to the Sun’s position across the entire vault of the sky, thus, to one that is able to perceive the direction of vibration, even a spot of blue sky can disclose the Sun’s position by its polarization patterns. Are bees endowed with this capacity?
To give further weight to the hypothesis that they were responding to polarization, Frisch performed an additional experiment :
The following test furnished an answer. The observation hive was set horizontally in a dark tent from which the dancers had a lateral view of a small area of blue sky. They danced correctly toward the west where their feeding place was located 200m away. When a round, rotatable polarizing foil was placed over the comb in a way as not to change the direction of the vibration of the polarized light from that part of the sky, they continued to dance correctly. If, however, I turned the foil right or left, the direction of the bees’ dance changed to the right or the left by corresponding angle values.
Von Frisch went on to conclude that for the bees, the sky revealed a pattern of polarized light from the Sun. He acknowledged that other creatures were known to see it, but that human beings and other vertebrates remained unendowed with this sense. We will revisit the admittedly more weak, but interesting case of the human ability to detect polarized light after exploring the visual world of some polarization-sensitive sea creatures, where this sense appears to be the most honed.
Cephalopods
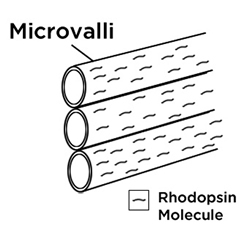
The cephalopods seem to share a relatively unique capability to respond to, and to reflect, patterns of polarized light. Cephalopods, with only one kind of squid as an exception, are colorblind, but their eyes serve them well, through an enhanced ability to selectively perceive linearly polarized light. The cephalopod eye has photoreceptors and corresponding hair-like microvilli which expand their surface area, and appear to be oriented orthogonally to adjacent ones, as seen in Figure 1b.
| FIGURE 1 | ||
 a) Close-up of cuttlefish eye |
b) Orthogonal microvilli |
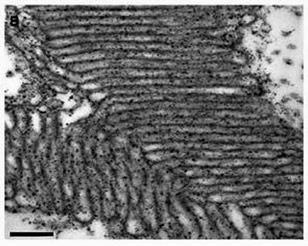
The common explanation for the polarization perception in cephalopods is that, since it is said that a specific population of retinal cells would be activated by polarized light in a specific plane, it is due to the orthogonal orientation of photoreceptors in the cephalopod, as in this case there would be a high population of retinal cells oriented in two different directions. It is said that human photoreceptors are less well organized, and that we humans barely perceive polarized light because the orientation of our visual pigment cells is “semi-random.” In the arthropods, and also the stomatopods which we will look at next, the visual pigments have a radial arrangement. Here is a common description of how polarized light interacts with visual pigments, which we will show to be rather too simple:
Visual pigment molecules are based on a single type of chromophore, whose highest absorption occurs when the molecule’s dipole is aligned with the e-vector axis of the light, making visual pigment molecules naturally polarization sensitive. In vertebrate rods and cones, the visual pigment is arranged in a semi-random array of axes, which makes the photoreceptor equally sensitive to any e-vector orientation when the light arrives parallel to the photoreceptor’s long axis.[8]
In this statement, there are a few problems which we should keep in mind. One, is that we don’t know exactly what causes the highest absorption of the light polarized in a given plane when aligned with the pigment, let alone how phototransduction occurs in eye, converting light into electrical signals. We know of the association, but polarization is a tricky phenomenon, because light itself is. But two, it would appear that it is the macro-organization of the pigments that matters, contrary to what this statement implies.
The so-called randomly organized pigments of the human eye do not appear to be as highly organized as they are in the eyes of these other creatures, if we consider the macro-organization of these pigments—but wouldn’t the same polarized light activate a portion of our retinal cells oriented in a parallel fashion, just not close-packed together, if the orientation of the pigments were the simple requirement? Perhaps the sheer number of pigments oriented in the same plane is simply not comparable to what the cephalopods have.
The fact that we do perceive some polarized light should mean that the organization of the pigments is not in fact random—assuming this has something to do with polarization sensitivity. This is besides the fact that claiming that any feature of human anatomy is semi-random, usually means something more like, “We don’t know how it is organized.” Accounts of the polarization sensitivity of humans, arthropods, stomotopods, and cephalopods all hinge on a particular arrangement of the visual pigments, but, as we have indicated, in each of these cases, each class represents a different arrangement.
Another paradox: The polarization-sensitive bees can perceive the colors white, yellow, blue, violet, and ultraviolet, but the polarization sensitivity of bees and other insects seems to correspond only to the ultraviolet wavelengths of light. But if the bees see five different colors, why would they only see polarized light in one of them? The simple radial arrangement of all visual pigments, as it is typically presented, does not account for this.
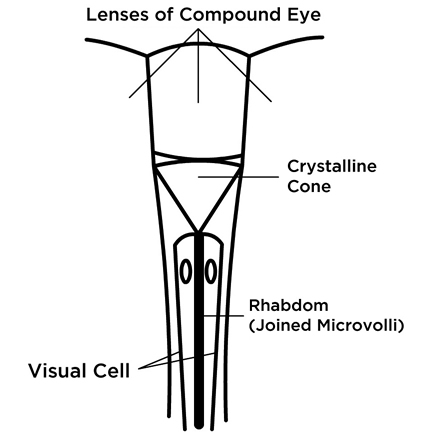
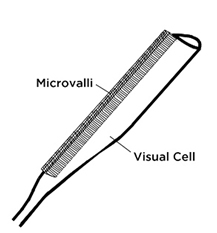
One explanation, is that in the region which is not sensitive to polarized light, there is a 180° rotation of the pigments along the length of the photoreceptor, canceling out the polarization. But even in the area receptive to UV light, there is a 40° rotation of the pigment.[9] This account does not quite match up with the descriptions of how polarized light interacts with visual pigments based on their perfect alignment, since high sensitivity to polarized light is apparently otherwise achieved with a 40° rotation of pigments through a twisting of the rhabdoms, a rod-shaped part of the insect’s eye (Figure 2). It seems as though the pigments in our human eye, though they be randomly arranged, should have an array of pigments spanning at least 40° in their orientation with respect to one another. But our polarized vision is clearly less acute, which means that this simple explanation of how “polarized vision” works doesn’t quite make it.
| FIGURE 2 | ||
 |
 |
|
 |
 |
|
|
LPAC/Chris Jadatz Several images of cuttlefish eye, from top left to bottom right with increasing resolution. |
||
In discussion of the polarization sensitivity of animals, there is heavy emphasis on the orientation of visual pigments, but this alone does not account for the phenomenon of polarization perception. It is not simply the organization of a substance which allows it to be sensitive to polarized light; the material itself determines the interaction with polarized light. It can also polarize light itself, in addition to simply being sensitive to it. In the human eye, it is thought that our ability to weakly perceive polarized light is additionally influenced by a crystal-like property of the cornea, which has its own slight polarizing effect on light. For the mantis shrimp, as we will see, the crystalline structure of the microvilli is said to affect the polarization. All we know is that the material and organization together seem to correspond to the ability to polarize light, and to selectively perceive it. The mechanism remains unclear, although it may have seemed somewhat intuitive at first; but it is the activity of the cephalopods and a handful of other creatures in response to the polarized light that we do know.
This capability has been tested more extensively with the cuttlefish, which has a camouflage capability that includes a polarization variable. This ability of the skin to polarize light seems to be especially prominent in the blue-green light range, a range in which the animal is colorblind. When placed in front of a blue, yellow, or a blue and yellow checkerboard background, the cuttlefish never changes its camouflage in response, when these colors are of the same intensity.[10] Despite lacking one aspect of a visual sense, they can respond to changes in the polarization of light much more efficiently than other creatures.
For example, a cuttlefish will respond differently to its own reflection, if seen through different polarized filters, and will change polarization patterns around other cuttlefish in displays of aggression or when attacking prey. Their skin demonstrates distinct patterns when seen though a polarized filter, which are otherwise not visible, and there are indications that the polarization of their skin is physiologically controlled. In one experiment, changing the chemical environment of the skin changed the polarization characteristic of the light reflected off of the skin. These examples indicate that cuttlefish may communicate with each other through induced polarization patterns in their skin, taken in addition to what it known about their orthogonal visual pigments.
The polarization is achieved through reflective cells called iridophores, which lie underneath a layer of chromatophore skin pigments. The chromatophores have small pigment sacs which expand, contract, and change shape to create the cuttlefish’s camouflage. The iridophores, or guanophores, are crystalline plates made of guanine, among other things, and are also used to produce colors in the cuttlefish’s camouflage. For example, purple can be created by a red chromatophore and an iridophore. The cuttlefish can also use an iridophore and a yellow chromatophore to produce a brighter green.
As for using these iridophores to create the polarization patterns, do the cuttlefish achieve this by the iridophores themselves changing in orientation with respect to the incident light, while being present over all of the skin? Or, are there special patterns of iridophores that have this induced polarization capability? In squid, it seems that the latter may be the case. But the change in polarization patterns is able to occur so quickly, that it is thought by researchers to be neurological (as opposed to hormonal), and researchers are currently puzzled as to how the changes in polarization can be induced within less than a second. Only very recently have nerve fibers been found in the vicinity of the iridophores. Prior to this, no squid had been known to have iridophores that are under neural control, and even this is still unproven, since the nerve fibers have only been found near the iridophores—no actual connection has yet been established.[11]
 The cuttlefish, in one of its camouflages. |
Another paradox about cuttlefish vision was communicated in a 2007 study which tested the “optomotor response” of cuttlefish, in response to moving patterns of contrasting stripes, and moving patterns of polarized stripes. In the optomotor response, the cuttlefish, in a tank surrounded by one of these backgrounds, should rotate around its center to follow the image that is circling around the tank. The cuttlefish did just this in response to the patterns of contrasting stripes (of different intensities), but not for polarized stripes. While this experiment was only done with one rare species of cuttlefish, it still puzzled researchers. The orthogonal structure of the eye’s pigments was present. Are polarization and intensity perceived differently by this cuttlefish, they asked? Are only certain kinds of visual cues involved in an OMR (Optical Mark Recognition) test? Or is the fish possibly not seeing polarized light, despite having the eye structure to account for it?[12]
How can insight into the control over the biological polarization mechanism, and the mechanism accounting for its perception, give us more insight into the still not-well-understood phenomenon of polarization? Is it achieved biologically by means which do not fully reconcile with our current explanations? This will be suggested even more in the case of the mantis shrimp.
An additional puzzling question for researchers is how the cuttlefish, who are colorblind, can match colors in their camouflage. They can perceive brightness and intensity, and patterns based on these contrasts, but how they are able to match colors, even in complete darkness, is puzzling to researchers. Using night-vision video, scientists at Woods Hole Marine Lab discovered that cuttlefish even match their background at night, when there isn’t enough light for color vision. Claims by some that this is explained by passively reflective cells called leucophores, do not seem to account for the sharp changes in patterns which they can induce. Dr. Roger Hanlon, who has written many research papers on the cuttlefish, and has done a lot of field work with them, was asked how the cuttlefish’s skin changes to any hue in the rainbow, although the animal has only one visual pigment which is sensitive to colored light at 492 nanometer (nm). He replied, “That’s a vexing question. We don’t know how it works.”[13] In the case of the cephalopods, we have a creature which discerns polarized light, and has the ability to induce changes in its skin polarization patterns in less than a second, by an unknown mechanism, which then appears to be seen by other cephalopods, who are colorblind, but can clearly perceive color in some way, as their camouflage demonstrates. Vision more generally seems to be quite perplexing!
However, even the polarized vision and communication capability of the cephalopods is not nearly as well developed as the capability of a specific kind of crustacean called the stomatopod, or mantis shrimp.
Mantis Shrimp
Not only does the mantis shrimp pack a serious punch, so fast that it can produce killer sonoluminescing bubbles, but these guys blow everyone out of the water in terms of a functional polarization-sensitive visual apparatus. The hyperspectral eyes of mantis shrimp, which perceive from the infrared to ultraviolet range (to 300 nm),[14] can also perceive linearly and circularly polarized light.
The tail of the male mantis shrimp, as well as other parts of their bodies, seem to emit circularly polarized light. When seen through a filter for either left-circularly polarized light or right-circularly polarized light, only one of these images of the tail will be illuminated.
It is unclear whether this is a kind of bioluminescence, a controlled reflection as in the case of the cephalopods, or simply the reflective nature of the material, although several articles imply that the males use this ability to “signal” others, implying that it is more than passive reflectivity.
The shrimp have 12 primary color pigments to our 3, and 4 which aid in polarization sensitivity. Each eye has three distinct parts, the two hemispheres and the midband, and is capable on its own of trinocular vision.
|
Mantis shrimp tail seen through left and right circularly polarized filters. |

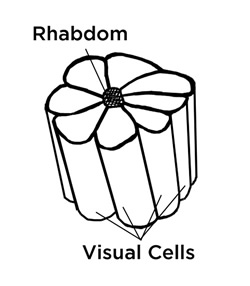
This midband is where most of the action occurs, being composed of many ommatidia, or “simple eyes,” each of which has long visual cells called rhabdoms, arranged and close-packed in a star pattern, pressing up against the ommatidia, similar to the insect eye. Here also, as in the cephalopods, we have tube-like microvilli, the light-sensitive part of the rhabdom, each of which points radially towards the center of the ommatidia, and which contains the pigment. Interestingly, more detailed studies reveal that the “small four lobed UV sensitive photoreceptor,” R-8, in the midband, is also said to be the one responsible for the circular polarization perception—two super-senses in one! “Circular polarization sensitivity is not innate to the R1-7 cells, but arises from the quarter-wave retardance of the overlying four-lobed R-8 cell.”[15] In some fish, and in bees and other insects, UV perception and polarization sensitivity are related, but only for linearly polarized light.
Seeing the circularly polarized light is thought to be unique to several species of mantis shrimp, although fireflies and scarab beetles can generate it; scarab beetles reflect it off of their liquid-crystal-like exoskeleton. One of several experiments used to detect the mantis shrimp’s sensitivity was done by giving them food with a flashing left circularly polarized light signal above it. Next to this station, would be a flashing right, circularly polarized light signal, but no food.[16] This would be repeated, and the positions of the two stations alternated, one with food, one without, but the light signals kept the same—the left circularly polarized light always at the station with food. When the food was removed, unbeknownst to the mantis shrimp, after having repeated this exercise many times, the mantis shrimp invariably went for the flashing left circularly polarized light signal. If this experiment were repeated with humans, our choice about which station to go to would have been arbitrary (or we might have a slight chance of making an informed guess, as we will soon see), whereas for the mantis shrimp, it would be informed by sensing some distinction between the left and right polarized light, though we don’t know exactly how this looks to them. How they sense the light is usually compared to the function of a quarter wave-plate, the non-biological mechanism we use to convert circularly polarized light to linearly polarized light. Accounting for the perceptive ability is based on this kind of quarter wave-plate being literally in the eye, in the R-8 cell overlaying the other rhabdom. It would be interesting to compare how the non-biological quarter-wave retarder in our labs is different from that in the mantis shrimp’s eye—and they do appear to be different. Is it a unique kind of crystalline structure, as is the case for our wave plates, which are made of calcite, quartz, or magnesium fluoride? It appears to be the case, but they are still quite different. The efficiency of their “wave plate” is said to be greater than even our own quarter-wave plates by a factor of 3. What accounts for this is unclear. As these researchers from Nature Photonics admit, the optical capabilities of the mantis shrimp’s eyes may be more advanced than some of our best noëtic instrumentation:
We have discovered a novel microvillar mechanism that acts as a remarkable achromatic optical device. Man-made retarders are among the most important and commonly used optical components, and the cellular structure we describe [of the mantis shrimp] significantly outperforms these current optics.[17]
This is aside from the fact that our optics are not also used to perceive linearly polarized UV light! The question of how alike, in fact, are the means by which humans with our instruments, and animals with their bodies, receive and produce polarized light, is forcefully posed by the case of the mantis shrimp. We can ask ourselves, what does the world look like to this creature? And although the circular polarized vision seems the most exotic, it is also interesting to ask how this creature perceives color. What does it look like to see based on the blending of 12 color pigments? Would you see “different colors,” or would variations be more striking? Would they blend differently? For a mantis shrimp, which colors would combine to make green? Blue and yellow, or completely different colors? What “color” is infrared or UV light for this creature? What does the visual field of an animal which can see all kinds of polarized light look like? Or what does it look like to have one eye with trinocular vision? Two? As we extend our concept of the sensorium, there seems to be a gap between the supposed impressions of these super-senses, their actual perceptions, and the actions of the creature, although these senses are not used for creativity. Some of what is unexplained lies within the “technology” itself. Although these creatures lack mind, untangling the problem of how these biological senses actually work, is a problem which continues to puzzle us, a problem of which these sea creatures, for example, are unaware, though they operate based on them to near perfection. Clearly a mantis shrimp and a human being do not see the same thing; the visual impressions received are thus not real objects, but different, contrasting impressions received from different sets of instruments. In the next case, we will show that our human visual map may have more resolution to it than we may assume from the most obvious impressions.
Humans and Haidinger’s Brush
After reviewing some cases of super polarized vision, the human capability to perceive polarized light may seem rather lame: a faint blue and yellow bow which you may or may not be able to see on your laptop screen or on a blue patch of sunny sky close to the horizon. You probably think that seeing circularly polarized light is out of the question—but seeing a diagonal brush which maintains its orientation as your head tilts, indicates that the light is circularly polarized, left or right, depending on the tilt! But surely this isn’t as useful as being able to communicate with other members of our species through secret polarization channels.
|
An exaggerated view of Haidinger’s brush. |

Haidinger’s brush is what is called an entoptic phenomena, and was discovered in 1844 by German physicist, geologist, and mineralogist Wilhelm Karl von Haidinger. Similar to the floaters you may see “on your eye,” Haidinger’s brush is also not something external. After all this discussion about the highly structured pigments in animal eyes, and our “practically random arrangement,” how is this faint polarization perception achieved? The matter is not completely settled. An article published in 2010 points out the flaws in a couple of theories, and posits the researchers’ own, which they tested through creating an artificial eye and camera. The reasons for being able to see the funny pattern of polarized light here also trace back to an organization of the eye’s pigments. But as you see in this brief summary, completely different models were said to be able to account for its perception. The 2010 study references previous theories:
Most models are based on either a possible radial or -tangential arrangement of absorbing elongated yellow pigments in the macula. Unfortunately, a radial alignment of anisotropically absorbing molecules along the nerve fibers which may be expected for highly elongated pigments would lead to reverse brush colors. Tangential alignment of the molecules orthogonally to the fibers would lead to the correct colors, but are unexpected and has never been experimentally observed.[18]
The researchers who wrote this critique say that they can produce the correct brush colors and orientation based on a particular cylindrical organization of a small population of blue cones in the fovea, a small section of the macula. They claim to have mimicked this organization in an artificial eye-like device, and say that they were able to photograph an image generated by this device which produced the blue brush when blue light was shown, and the yellow brush when red and green light was shown. However, accounts of people seem to indicate that the brush is not perceived at all with red light, but that specifically blue light is required. The cause of the particular faint colors of the brushes is not clearly related causally to the color of the perceived light, nor how or whether it depends on the organization of the eye’s pigments—for example, radial or tangential, as implied above, where both could be used to explain the perception. Does it depend on yellow or blue pigments? Both explanations have been given. Another account suggests that it may be a birefringence in the eye itself which accounts for the particular colors. As we can see, there are and have been many theories put forward. Many models claim to account for some aspect of the perception, but none claim to have reproduced it completely in the same way as the human eye does.
It is nonetheless interesting that this last model should rely on a specific arrangement of the eye’s blue cones, which are relatively sparse in the human eye anyway— only 2% of our cones are blue cones, but they are highly sensitive for yet unexplained reasons. Most of us would not consider ourselves to be blue colorblind, despite having so few blue cones. In the area of the fovea, the percentage of blue cones is even less than 2%. Blue light has proved important for other phenomena referenced in this report, including bird magnetoreception, etiolation, and certain biological rhythms. But at least one account claims that the fovea is too small to account for the perception based on the size of the brush.
The above apparatus as described, a simple machine involving not much more than a lens, a glass cylinder, and a “blue mosaic on a screen,” cannot be seriously treated as an analogue to the human eye. And also, the unique arrangement of a very small number of blue cones, which this model relied on, does not, on the surface, account for other phenomena associated with Haidinger’s brush. You may want to understand why you can see Haidinger’s brush, or why you can’t, because it may have to do with your overall visual health, as researchers are finding out.
The ability to sharply perceive Haidinger’s brush in a particular eye, has been linked in many people to the “dominant eye,” which is also a puzzling phenomenon. But those functions which we associate with eye dominance, dealing with perception much more generally, do not on the surface to account for why the dominant eye would be able to perceive a sharper Haidinger’s brush. Apparently, the ability or lack thereof to see Haidinger’s brush is even used to diagnose some degenerative conditions in the eye:
The absence of a photographically visible polarization pattern is an indication of macular dysfunction due to senior macular degeneration, angioid streaks, or diabetic retinoplasty, and thus the phenomenon can be useful for diagnosing diseases affecting the macula.... Perception of Haidinger’s brushes may indicate a healthy eye, and the inability of perception of these brushes indicates certain visual dysfunctions.[19]
Additionally, patients with certain kinds of strabismus, or “turning eye,” can be trained to view objects with the correct part of their eye by lining up the Haidinger’s brush with the object they are trying to look at.
We have, with Haidinger’s brush, a perception much less stark than those used by the bee, cephalopod, or mantis shrimp to function day to day, but which may be just that significant for our own vision all the time, despite the fact that we aren’t consciously seeing it all the time. However useful the ability to perceive Haidinger’s brush may be for making the above diagnoses, it is only correlated with these various degenerative eye conditions—there is not a demonstrable causal connection between the them.
Perhaps we could refer to it as a kind of visual “weak force.” That is, something barely perceived or sensed by us, as, for example, in the case of various low-intensity kinds of radiation which play some critical role in the optimal functioning of an organism. Here, we have a faint, or low-intensity perception, which seems to play some more critical role for the function of vision. Perhaps the true cause for it would redefine our notion of vision itself—but with various and completely different models claiming to explain it, we are not there yet.
Let us, as Bernhard Riemann did for the investigation of the ear, start our investigations of vision based on taking into account what the animal and human visual apparatuses do, and allow that to shake up our models of how vision must function.[20] It does seem to be clear, that based on the function of the human eye generally (and the very intentional role of human beings!), claims that anything about its organization are random, as compared to the eyes of animals, seem more dishonest than anything. Such statements should be reformulated to state that we don’t fully understand the reasons behind the particular organization of the eye. Then again, the eye itself exists and functions based on its own relationship to cosmic radiation, polarized, unpolarized, and of varying intensities. Is there a cause which lies completely outside the domain of the rods and cones of the eye, as might also be the case for distinct closed-eye visual noise, colors, and patterns, or those you see when pressing or rubbing a closed eye? Or the lights seen by numerous astronauts, which appear when they close their eyes?[21] In addition, auroral “hearing,” bird magnetoreception, the phenomenon of synesthesia, and the case of someone like Helen Keller, can all cause us to wonder if there is not more to vision as a sense, than we might have assumed from the most obvious impressions.[22]
And despite the greater intensity and clear utility of the animal polarization sense, our own seemingly weaker visual perceptions do not leave us weaker as a species. But, could we further increase our power over nature through honing our own polarization sense, through our man-made instruments and even our own biological instrument? Based on how much time most of us spend on a given day staring at an LCD screen on our laptop, it may be that we have been subconsciously training ourselves to block out the perception of Haidinger’s brush, as a kind of unwanted visual background noise. For Vikings who navigated the seas using pieces of Iceland spar to locate the Sun on a cloudy day, the polarization sense was second nature, and a matter of survival. Perhaps some of them were unaware that it was polarization which they were responding to, as you yourself might have been unaware of what generated the faint perception we have now identified as the human biological polarization sense. What other kinds of weak impressions, or phenomena more generally, could you be responding to, unknowingly?
[1]. See accompanying piece by Jason Ross, “What Is Circularly Polarized Light?” Also see video, Louis Pasteur: the Science of Life,” on the LaRouche PAC website.
[2]http://www.livescience.com/history/070302_viking_navigation.html. or http://www.polarization.com, a very useful website for this and other references from this report.
[3]. Polarized vision of many insects, such as dragonflies, can be deadly when they are tricked into laying their eggs on murderous solar panels, which they mistake for water because of the reflected polarized light. Let us be rid of these killer panels!
[4]. As Karl von Frisch said of the bees, “thus the language of the bee, which was initially brought to our attention by the physiology of sense perception ... led to general questions of orientation in time and space.” See Peter Martinson, “Following the Beat of a Different Drummer,” Ben Deniston, “Magnetoreception,” and Oyang Teng, “Insects and Infrared,” in this issue.
[5]. The polarization of the Sun’s light is greatest 90° from the Sun, something you can test with polarized sunglasses. If the bee can so precisely indicate the location of the food based on this kind of reading, it is not hard to imagine that the polarization pattern seen by the bees has more resolution than this.
[6]. http://nobelprize.org/nobel_prizes/medicine/laureates/1973/frisch-lecture.pdf
[7]. Von Frisch alludes to polarization in a particular color of light, an indication that the eyes’ pigments themselves are contributing to the polarization sensitivity. As it turns out, bees, and many insects, perceive polarized light distinctly in the UV range. Other experiments have shown that the perception of polarized light by bees can still be somewhat efficient in a partially cloudy sky, which would support this idea. There are conflicting accounts about whether bees see polarized blue light, to which von Frisch alluded. We will further examine what this means and how it is determined a bit later in this report.
[8]. Lydia M. Mathger, Nadav Shashar, Roger T. Hanlon, “Do Cephalopods Communicate Using Polarized Light Reflections from Their Skin?” Journal of Experimental Biology, 212, 2,133-2,140, Doi:10.1242/jeb.020800 (2009).
[9]. To what extent this is based on observation, or just a model, was not clear from the account.
[10]. Lydia M. Mathger et al., “Color blindness and contrast perception in cuttlefish determined by a visual sensorimotor assay,” Vision Research, 46 (2006) 1,746-1,753, doi: 10.1016/j.vires.2005.09.035
[11]. Nadav Shashar et al., “Polarization reflecting iridophores in the arms of the squid Loligo pealeii,” Biological Bulletin, 201:267-268 (2001).
[12]. Nadav Shashar and Anne-Sophie Darmaillacq, “Lack of polarization optomotor response in the cuttlefish Sepia elongata,” Physiology & Behavior (2008), doi:10.1016/j.physbeh.2008.01.018
[13]. http://www.nytimes.com/2008/02/19/science/19camo.html
[14]. For cell perception of UV below this frequency, see Cody Jones, “Cosmic Bio-Radiation: Casting Gurwitsch in the Light of Vernadsky,” http://www.larouchepac.com/files/CodyJones-BioCosmo_0.pdf
[15]. S. Kleinlogel and A.G. White, “The Secret World of Shrimps: Polarization Vision at Its Best.” PloS ONE 3(5): e2190. Doi:10.1371/journal.pone.0002190 (2008).
[16]. “How Mantis Shrimp see circularly polarized light,” Aug. 16, 2010, http://arthropoda.southernfriedscience.com/?p=2964
[17]. N.W. Roberts, T-H Chiou, M.J. Marshall, T.W. Cronin, “A biological quarter wave retarder with excellent achromaticity in the visible wave length region,” Nature Photonics, Doi:10.1038/NPhoton/2009.189 (2009).
[18]. Albert Le Floch, Guy Ropars, et al. “The polarization sense in human vision,” Vision Research, 50, 2,048-2,054, doi:10.1016/j.visres.2010.07.007 (2010).
[19]. Horvath Gabor and Varju Dezso, “Polarized Light in Animal Vision: Polarization Patterns in Nature,” http://www.uni-tuebingen.de/cog/literature/literatur-Dateien/2003/HoVa_bookcontensts03.pdf
[20]. See Aaron Halevy on Riemann’s approach to hearing, “The Sounds of a Cosmic Chorus,” this issue, and, http://www.larouchepac.com/files/AaronHalevy-CosmicChorus_0.pdf
[21]. http://www.space.com/scienceastronomy/mir_lights_030416.html
[22]. See Oyang Teng, “Synesthesia: Beyond the Five Senses,” and Sky Shields, “Unheard Melodies: Electric and Magnetic Senses in Humans,” in this issue.